Pupil Dilation during Reward Anticipation Is Correlated to Depressive Symptom Load in Patients with Major Depressive Disorder

 ,
,
Abstract
:1. Introduction
2. Method
2.1. Subjects
2.2. Psychometric Data
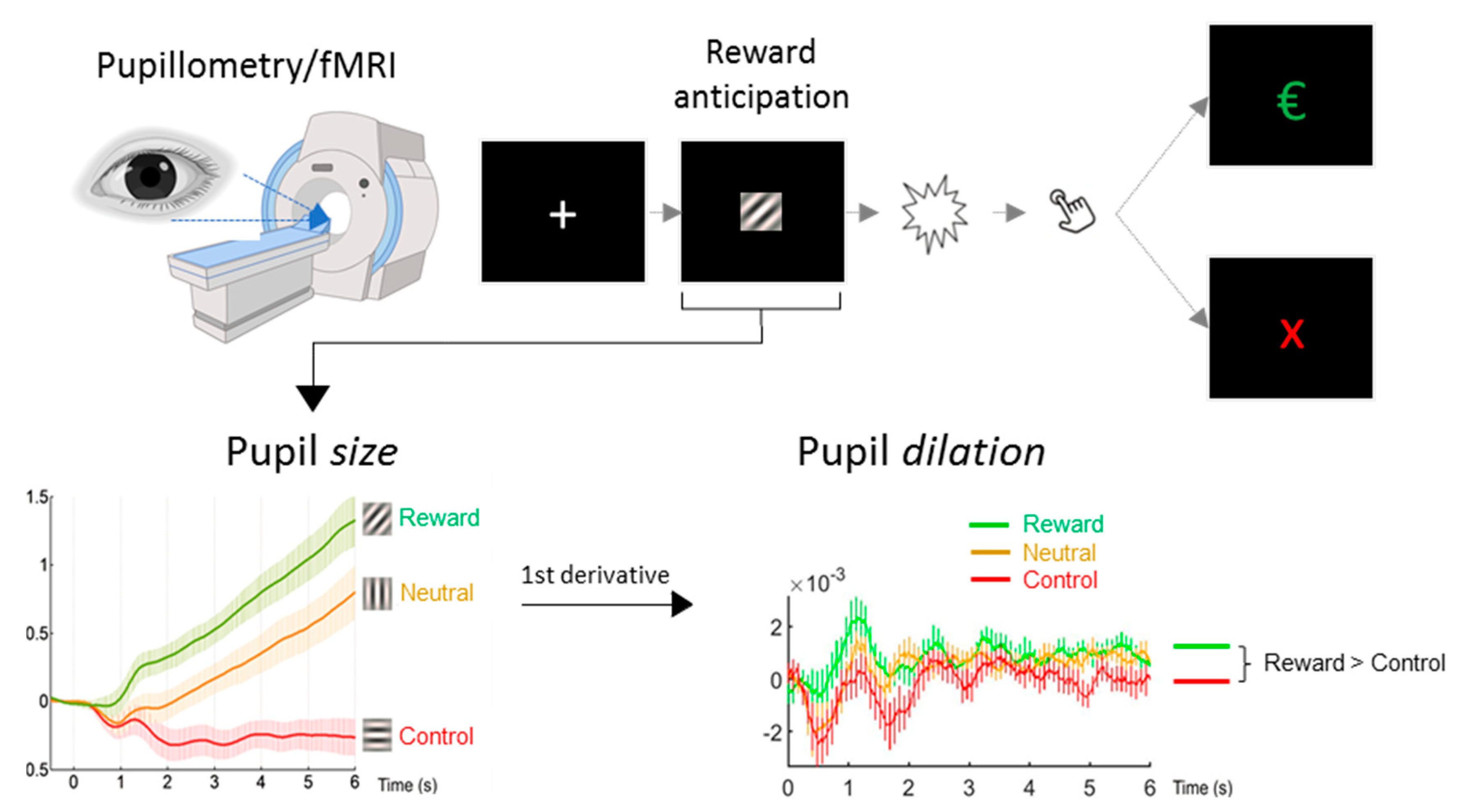
2.3. Paradigm
2.4. Pupillometry
2.5. fMRI
2.6. Statistical Analyses
3. Results
3.1. Classical Group Comparison
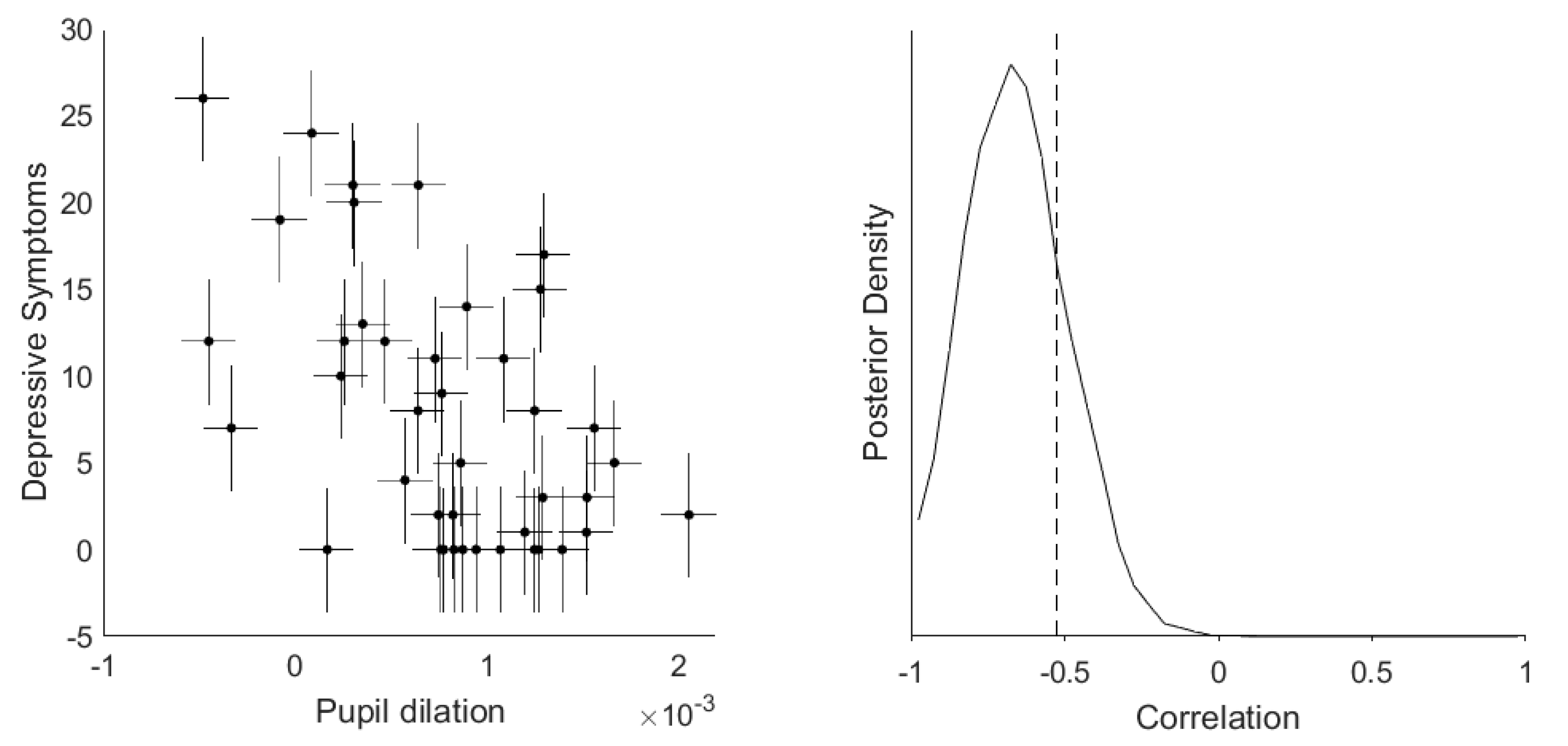
3.2. Dimensional Correlation Analyses between Depressive Symptoms and Pupil Dilation
3.3. Correlation between Pupil Dilation and Motor Response
3.4. FMRI Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Depression and Other Common Mental Disorders: Global Health Estimates; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Russo, S.J.; Nestler, E.J. The brain reward circuitry in mood disorders. Nat. Rev. Neurosci. 2013, 14, 609. [Google Scholar] [CrossRef] [Green Version]
- Nestler, E.J.; Carlezon, W.A. The mesolimbic dopamine reward circuit in depression. Biol. Psychiatry 2006, 59, 1151–1159. [Google Scholar] [CrossRef]
- Whitton, A.E.; Treadway, M.T.; Pizzagalli, D.A. Reward processing dysfunction in major depression, bipolar disorder and schizophrenia. Curr. Opin. Psychiatry 2015, 28, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Kupfer, D.J.; Frank, E.; Phillips, M.L. Major depressive disorder: New clinical, neurobiological, and treatment perspectives. Lancet 2012, 379, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Gradin, V.B.; Kumar, P.; Waiter, G.; Ahearn, T.; Stickle, C.; Milders, M.; Reid, I.; Hall, J.; Steele, J.D. Expected value and prediction error abnormalities in depression and schizophrenia. Brain J. Neurol. 2011, 134, 1751–1764. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Goer, F.; Murray, L.; Dillon, D.G.; Beltzer, M.L.; Cohen, A.L.; Brooks, N.H.; Pizzagalli, D.A. Impaired reward prediction error encoding and striatal-midbrain connectivity in depression. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2018, 43, 1581–1588. [Google Scholar] [CrossRef]
- Kumar, P.; Waiter, G.; Ahearn, T.; Milders, M.; Reid, I.; Steele, J.D. Abnormal temporal difference reward-learning signals in major depression. Brain J. Neurol. 2008, 131, 2084–2093. [Google Scholar] [CrossRef] [Green Version]
- Rutledge, R.B.; Moutoussis, M.; Smittenaar, P.; Zeidman, P.; Taylor, T.; Hrynkiewicz, L.; Lam, J.; Skandali, N.; Siegel, J.Z.; Ousdal, O.T.; et al. Association of neural and emotional impacts of reward prediction errors with major depression. JAMA Psychiatry 2017, 74, 790–797. [Google Scholar] [CrossRef]
- Rudebeck, P.H.; Putnam, P.T.; Daniels, T.E.; Yang, T.; Mitz, A.R.; Rhodes, S.E.V.; Murray, E.A. A role for primate subgenual cingulate cortex in sustaining autonomic arousal. Proc. Natl. Acad. Sci. USA 2014, 111, 5391–5396. [Google Scholar] [CrossRef] [Green Version]
- Aston-Jones, G.; Cohen, J.D. An integrative theory of locus coeruleus-norepinephrine function: Adaptive gain and optimal performance. Annu. Rev. Neurosci. 2005, 28, 403–450. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.; Leuchs, L.; Czisch, M.; Samann, P.G.; Spoormaker, V.I. Disentangling reward anticipation with simultaneous pupillometry/fMRI. NeuroImage 2018, 178, 11–22. [Google Scholar] [CrossRef]
- Knutson, B.; Fong, G.W.; Adams, C.M.; Varner, J.L.; Hommer, D. Dissociation of reward anticipation and outcome with event-related fMRI. Neuroreport 2001, 12, 3683–3687. [Google Scholar] [CrossRef] [Green Version]
- Bruckl, T.M.; Spoormaker, V.I.; Samann, P.G.; Brem, A.K.; Henco, L.; Czamara, D.; Elbau, I.; Grandi, N.C.; Jollans, L.; Kuhnel, A.; et al. The biological classification of mental disorders (BeCOME) study: A protocol for an observational deep-phenotyping study for the identification of biological subtypes. BMC Psychiatry 2020, 20, 213. [Google Scholar] [CrossRef]
- Wittchen, H.U.; Lachner, G.; Wunderlich, U.; Pfister, H. Test-retest reliability of the computerized DSM-IV version of the Munich-Composite International Diagnostic Interview (M-CIDI). Soc. Psychiatry Psychiatr. Epidemiol. 1998, 33, 568–578. [Google Scholar] [CrossRef]
- Kuhner, C.; Burger, C.; Keller, F.; Hautzinger, M. Reliability and validity of the revised beck depression inventory (BDI-II). Results from German samples. Der Nervenarzt 2007, 78, 651–656. [Google Scholar] [CrossRef]
- Munafò, M.R.; Nosek, B.A.; Bishop, D.V.M.; Button, K.S.; Chambers, C.D.; Percie du Sert, N.; Simonsohn, U.; Wagenmakers, E.-J.; Ware, J.J.; Ioannidis, J.P.A. A manifesto for reproducible science. Nat. Hum. Behav. 2017, 1, 0021. [Google Scholar] [CrossRef] [Green Version]
- Wagenmakers, E.-J.; Love, J.; Marsman, M.; Jamil, T.; Ly, A.; Verhagen, J.; Selker, R.; Gronau, Q.F.; Dropmann, D.; Boutin, B.; et al. Bayesian inference for psychology. Part II: Example applications with JASP. Psychon. Bull. Rev. 2018, 25, 58–76. [Google Scholar] [CrossRef] [Green Version]
- Jeffreys, H. Theory of Probability, 3rd ed.; Oxford University Press: Oxford, UK, 1961. [Google Scholar]
- Lee, M.D.; Wagenmakers, E.-J. Bayesian Cognitive Modeling: A Practical Course; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Matzke, D.; Ly, A.; Selker, R.; Weeda, W.D.; Scheibehenne, B.; Lee, M.D.; Wagenmakers, E.-J. Bayesian inference for correlations in the presence of measurement error and estimation uncertainty. Collabra Psychol. 2017, 3, 25. [Google Scholar] [CrossRef]
- Nassar, M.R.; Rumsey, K.M.; Wilson, R.C.; Parikh, K.; Heasly, B.; Gold, J.I. Rational regulation of learning dynamics by pupil-linked arousal systems. Nat. Neurosci. 2012, 15, 1040. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Park, J. Using SPM 12’s second-level bayesian inference procedure for fMRI analysis: Practical guidelines for end users. Front. Neuroinform. 2018, 12, 1. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, S.J.; Pizzagalli, D.A.; Sproule, B.A.; Kennedy, S.H. Assessing anhedonia in depression: Potentials and pitfalls. Neurosci. Biobehav. Rev. 2016, 65, 21–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzagalli, D.A. Depression, stress, and anhedonia: Toward a synthesis and integrated model. Annu. Rev. Clin. Psychol. 2014, 10, 393–423. [Google Scholar] [CrossRef] [Green Version]
- Starcke, K.; Brand, M. Decision making under stress: A selective review. Neurosci. Biobehav. Rev. 2012, 36, 1228–1248. [Google Scholar] [CrossRef] [PubMed]
- Huys, Q.J.; Pizzagalli, D.A.; Bogdan, R.; Dayan, P. Mapping anhedonia onto reinforcement learning: A behavioural meta-analysis. Biol. Mood Anxiety Disord. 2013, 3, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huys, Q.J.M.; Gölzer, M.; Friedel, E.; Heinz, A.; Cools, R.; Dayan, P.; Dolan, R.J. The specificity of Pavlovian regulation is associated with recovery from depression. Psychol. Med. 2016, 46, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Sharp, C.; Kim, S.; Herman, L.; Pane, H.; Reuter, T.; Strathearn, L. Major depression in mothers predicts reduced ventral striatum activation in adolescent female offspring with and without depression. J. Abnorm. Psychol. 2014, 123, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Satterthwaite, T.D.; Kable, J.W.; Vandekar, L.; Katchmar, N.; Bassett, D.S.; Baldassano, C.F.; Ruparel, K.; Elliott, M.A.; Sheline, Y.I.; Gur, R.C.; et al. Common and dissociable dysfunction of the reward system in bipolar and unipolar depression. Neuropsychopharmacology 2015, 40, 2258–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diego, A.; Pizzagalli, P.D.; Avram, J.; Holmes, A.M.; Daniel, G.; Dillon, P.D.; Elena, L.; Goetz, B.A.; Jeffrey, L.; Birk, B.A.; et al. Reduced caudate and nucleus accumbens response to rewards in unmedicated individuals with major depressive disorder. Am. J. Psychiat. 2009, 166, 702–710. [Google Scholar] [CrossRef] [Green Version]
- Keren, H.; O’Callaghan, G.; Vidal-Ribas, P.; George, A.B.; Melissa, A.B.; Leibenluft, E.; Pedro, M.P.; Meffert, L.; Kaiser, A.; Wolke, S.; et al. Reward processing in depression: A conceptual and meta-analytic review across fMRI and EEG studies. Am. J. Psychiatry 2018, 175, 1111–1120. [Google Scholar] [CrossRef] [Green Version]
- Oei, N.Y.L.; Both, S.; van Heemst, D.; van der Grond, J. Acute stress-induced cortisol elevations mediate reward system activity during subconscious processing of sexual stimuli. Psychoneuroendocrinology 2014, 39, 111–120. [Google Scholar] [CrossRef]
- Kumar, P.; Berghorst, L.H.; Nickerson, L.D.; Dutra, S.J.; Goer, F.K.; Greve, D.N.; Pizzagalli, D.A. Differential effects of acute stress on anticipatory and consummatory phases of reward processing. Neuroscience 2014, 266, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Plichta, M.M.; Schwarz, A.J.; Grimm, O.; Morgen, K.; Mier, D.; Haddad, L.; Gerdes, A.B.M.; Sauer, C.; Tost, H.; Esslinger, C.; et al. Test–retest reliability of evoked BOLD signals from a cognitive–emotive fMRI test battery. NeuroImage 2012, 60, 1746–1758. [Google Scholar] [CrossRef] [Green Version]
- McTeague, L.M.; Rosenberg, B.M.; Lopez, J.W.; Carreon, D.M.; Huemer, J.; Jiang, Y.; Chick, C.F.; Eickhoff, S.B.; Etkin, A. Identification of common neural circuit disruptions in emotional processing across psychiatric disorders. Am. J. Psychiatry 2020, 177, 411–421. [Google Scholar] [CrossRef]
- Bartova, L.; Meyer, B.M.; Diers, K.; Rabl, U.; Scharinger, C.; Popovic, A.; Pail, G.; Kalcher, K.; Boubela, R.N.; Huemer, J.; et al. Reduced default mode network suppression during a working memory task in remitted major depression. J. Psychiatr. Res. 2015, 64, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Sheline, Y.I.; Barch, D.M.; Price, J.L.; Rundle, M.M.; Vaishnavi, S.N.; Snyder, A.Z.; Mintun, M.A.; Wang, S.; Coalson, R.S.; Raichle, M.E. The default mode network and self-referential processes in depression. Proc. Natl. Acad. Sci. USA 2009, 106, 1942–1947. [Google Scholar] [CrossRef] [Green Version]
- Jones, N.P.; Siegle, G.J.; Muelly, E.R.; Haggerty, A.; Ghinassi, F. Poor performance on cognitive tasks in depression: Doing too much or not enough? Cogn. Affect. Behav. Neurosci. 2010, 10, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Jones, N.P.; Siegle, G.J.; Mandell, D. Motivational and emotional influences on cognitive control in depression: A pupillometry study. Cogn. Affect. Behav. Neurosci. 2015, 15, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Siegle, G.J.; Steinhauer, S.R.; Thase, M.E. Pupillary assessment and computational modeling of the Stroop task in depression. Int. J. Psychophysiol. Off. J. Int. Organ. Psychophysiol. 2004, 52, 63–76. [Google Scholar] [CrossRef]
- Siegle, G.J.; Granholm, E.; Ingram, R.E.; Matt, G.E. Pupillary and reaction time measures of sustained processing of negative information in depression. Biol. Psychiatry 2001, 49, 624–636. [Google Scholar] [CrossRef]
- Silk, J.S.; Dahl, R.E.; Ryan, N.D.; Forbes, E.E.; Axelson, D.A.; Birmaher, B.; Siegle, G.J. Pupillary reactivity to emotional information in child and adolescent depression: Links to clinical and ecological measures. Am. J. Psychiatry 2007, 164, 1873–1880. [Google Scholar] [CrossRef]
- Steidtmann, D.; Ingram, R.E.; Siegle, G.J. Pupil response to negative emotional information in individuals at risk for depression. Cogn. Emot. 2010, 24, 480–496. [Google Scholar] [CrossRef]
- Burkhouse, K.L.; Siegle, G.J.; Woody, M.L.; Kudinova, A.Y.; Gibb, B.E. Pupillary reactivity to sad stimuli as a biomarker of depression risk: Evidence from a prospective study of children. J. Abnorm. Psychol. 2015, 124, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.P.; Siegle, G.J.; Proud, L.; Silk, J.S.; Hardy, D.; Keljo, D.J.; Dahl, R.E.; Szigethy, E. Impact of inflammatory bowel disease and high-dose steroid exposure on pupillary responses to negative information in pediatric depression. Psychosom. Med. 2011, 73, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegle, G.J.; Steinhauer, S.R.; Friedman, E.S.; Thompson, W.S.; Thase, M.E. Remission prognosis for cognitive therapy for recurrent depression using the pupil: Utility and neural correlates. Biol. Psychiatry 2011, 69, 726–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schumann, A.; Andrack, C.; Bar, K.J. Differences of sympathetic and parasympathetic modulation in major depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 79, 324–331. [Google Scholar] [CrossRef]
- Sekaninova, N.; Ondrejka, I.; Bona Olexova, L.; Visnovcova, Z.; Mestanik, M.; Cesnekova, D.; Hamrakova, A.; Tonhajzerova, I. Oculometric behavior assessed by pupil response is altered in adolescent depression. Physiol. Res. 2019, 68, S325–S338. [Google Scholar] [CrossRef]
- Mestanikova, A.; Ondrejka, I.; Mestanik, M.; Cesnekova, D.; Visnovcova, Z.; Bujnakova, I.; Oppa, M.; Calkovska, A.; Tonhajzerova, I. Pupillary light reflex is altered in adolescent depression. Physiol. Res. 2017, 66, S277–S284. [Google Scholar] [CrossRef]
- Wang, J.; Fan, Y.; Zhao, X.; Chen, N. Pupillometry in Chinese female patients with depression: A pilot study. Int. J. Environ. Res. Public Health 2014, 11, 2236–2243. [Google Scholar] [CrossRef] [Green Version]
- Bar, K.J.; Greiner, W.; Jochum, T.; Friedrich, M.; Wagner, G.; Sauer, H. The influence of major depression and its treatment on heart rate variability and pupillary light reflex parameters. J. Affect. Disord. 2004, 82, 245–252. [Google Scholar] [CrossRef]
- Murphy, P.R.; O’Connell, R.G.; O’Sullivan, M.; Robertson, I.H.; Balsters, J.H. Pupil diameter covaries with BOLD activity in human locus coeruleus. Hum. Brain Mapp. 2014, 35, 4140–4154. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.; Li, Y.; Kalwani, R.M.; Gold, J.I. Relationships between pupil diameter and neuronal activity in the locus coeruleus, colliculi, and cingulate cortex. Neuron 2016, 89, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Reimer, J.; McGinley, M.J.; Liu, Y.; Rodenkirch, C.; Wang, Q.; McCormick, D.A.; Tolias, A.S. Pupil fluctuations track rapid changes in adrenergic and cholinergic activity in cortex. Nat. Commun. 2016, 7, 13289. [Google Scholar] [CrossRef]
- Larsen, R.S.; Waters, J. Neuromodulatory correlates of pupil dilation. Front. Neural Circuits 2018, 12, 21. [Google Scholar] [CrossRef] [Green Version]
- Schmid, Y.; Enzler, F.; Gasser, P.; Grouzmann, E.; Preller, K.H.; Vollenweider, F.X.; Brenneisen, R.; Muller, F.; Borgwardt, S.; Liechti, M.E. Acute effects of lysergic acid diethylamide in healthy subjects. Biol. Psychiatry 2015, 78, 544–553. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Ramage, A.G.; Koss, M.C. Pharmacological studies of 8-OH-DPAT-induced pupillary dilation in anesthetized rats. Eur. J. Pharmacol. 2004, 489, 207–213. [Google Scholar] [CrossRef]
- Einhauser, W.; Stout, J.; Koch, C.; Carter, O. Pupil dilation reflects perceptual selection and predicts subsequent stability in perceptual rivalry. Proc. Natl. Acad. Sci. USA 2008, 105, 1704–1709. [Google Scholar] [CrossRef] [Green Version]
- Insel, T.; Cuthbert, B.; Garvey, M.; Heinssen, R.; Pine, D.S.; Quinn, K.; Sanislow, C.; Wang, P. Research domain criteria (RDoC): Toward a new classification framework for research on mental disorders. Am. J. Psychiatry 2010, 167, 748–751. [Google Scholar] [CrossRef] [Green Version]
- Schumann, G.; Binder, E.B.; Holte, A.; de Kloet, E.R.; Oedegaard, K.J.; Robbins, T.W.; Walker-Tilley, T.R.; Bitter, I.; Brown, V.J.; Buitelaar, J.; et al. Stratified medicine for mental disorders. Eur. Neuropsychopharmacol. 2014, 24, 5–50. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 12-Month | Lifetime | |||
|---|---|---|---|---|
| DSM-IV Diagnoses | n | % | n | % |
| Any anxiety disorder | 22 | 53.6 | 21 | 51.2 |
| Panic disorder | 5 | 12.2 | 3 | 7.3 |
| Specific phobia | 17 | 41.5 | 5 | 12.2 |
| Generalized Anxiety Disorder | 0 | 0 | 11 | 26.8 |
| Posttraumatic Stress Disorder | 2 | 4.9 | 4 | 9.8 |
| Any substance use disorder | 3 | 7.3 | 10 | 24.4 |
| Any comorbid diagnosis | 9 | 22.0 | 20 | 48.8 |
| Just one comorbid diagnosis | 6 | 14.6 | 10 | 24.4 |
| Two comorbid diagnosis | 3 | 7.3 | 6 | 14.6 |
| more than two comorbid diagnoses | 0 | 0 | 4 | 9.8 |
| Comorbid with just an anxiety disorder | 0 | 0 | 3 | 7.3 |
| Comorbid with an anxiety and a substance use disorder | 0 | 0 | 3 | 7.3 |
| Region Name | MNI Coordinates | Cluster Extent | Peak Log Odds |
|---|---|---|---|
| Contrast [reward > nonresponse control] | |||
| Dorsal anterior cingulate | [8 14 40] | 3013 | 36.0 |
| Insula (L) | [−36 16 −6] | 2724 | 36.0 |
| Insula (R) | [36 20 −10] | 2572 | 36.0 |
| Ventral striatum (L) | [−8 8 0] | 1337 | 36.0 |
| Ventral striatum (R) | [10 6 −2] | „ | „ |
| Contrast [reward < nonresponse control] | |||
| Posterior cingulate | [10 −56 20] | 1848 | 34.7 |
| Medial prefrontal cortex | [−6 56 0] | 786 | 22.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schneider, M.; Elbau, I.G.; Nantawisarakul, T.; Pöhlchen, D.; Brückl, T.; BeCOME Working Group; Czisch, M.; Saemann, P.G.; Lee, M.D.; Binder, E.B.; et al. Pupil Dilation during Reward Anticipation Is Correlated to Depressive Symptom Load in Patients with Major Depressive Disorder. Brain Sci. 2020, 10, 906. https://doi.org/10.3390/brainsci10120906
Schneider M, Elbau IG, Nantawisarakul T, Pöhlchen D, Brückl T, BeCOME Working Group, Czisch M, Saemann PG, Lee MD, Binder EB, et al. Pupil Dilation during Reward Anticipation Is Correlated to Depressive Symptom Load in Patients with Major Depressive Disorder. Brain Sciences. 2020; 10(12):906. https://doi.org/10.3390/brainsci10120906
Chicago/Turabian StyleSchneider, Max, Immanuel G. Elbau, Teachawidd Nantawisarakul, Dorothee Pöhlchen, Tanja Brückl, BeCOME Working Group, Michael Czisch, Philipp G. Saemann, Michael D. Lee, Elisabeth B. Binder, and et al. 2020. "Pupil Dilation during Reward Anticipation Is Correlated to Depressive Symptom Load in Patients with Major Depressive Disorder" Brain Sciences 10, no. 12: 906. https://doi.org/10.3390/brainsci10120906